Dark Fluid DNA
Dark matter fluid dynamics explains covalent bond forces between atoms, and affects how amino acids bond to form protein chains.
There are 61 codons that are used to assemble the 20 different types of amino acids. Research focuses on the the amino acids in the protein chains. Despite the fact that it’s been demonstrated that “synonymous codons” cannot be simply replaced, not much research has gone into whether the codon might influence how the amino acid bonds. The theory is that the codon influences the speed of assembly and that influences how the protein folds.
Unlike the traditional theory of covalent bonds though, orbital signatures have angular momentum that will be conserved. The signature of this angular momentum waveform has information, history.
With dark fluid dynamics, I’ll show how the codon’s orbital signature is transferred to the amino acid using the tRNA. This magnetic fluid attraction affects how the amino acid bonds with both others and RNA nucleotides.
The Evidence
This is an evolving discovery, a part time project but see what I’ve found in a short time:
- Ribosomal properties map onto its rRNA.
- Codon patterns map to Alphafold’s “high confidence” features in the protein OvalY of Gallus Gallus (the chicken).
- Codon patterns where tp53 bonds with other copies of itself.
It appears that cellular dynamics are not based on chaos and entropy but instead programmed by fluid flow attraction and repulsion patterns coded into proteins and rRNA.
If you don’t believe me, try mapping out a few sequences yourself?

How does this work? Since dark matter fluid is the force behind magnetism, let’s use that analogy. Did you know that a stronger magnet realigns a weaker one without losing energy itself?
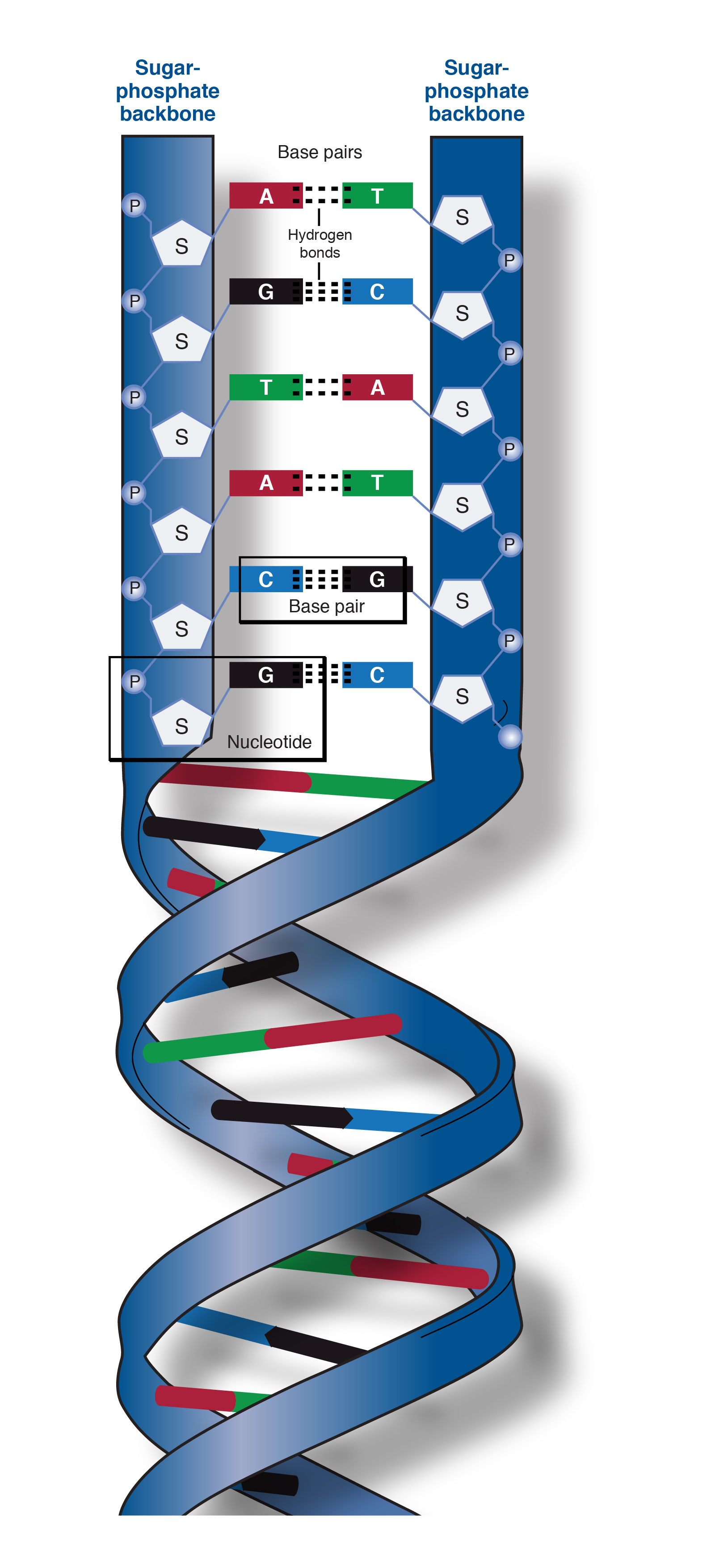
With that lens, opposite base pairs in DNA are recharging each other’s orbital signatures all the time while holding each other in place. Opposite pairs form a covalent bond from the opposite signatures they create and maintain. They recycle figure-8 dark fluid, aligning energy to minimize turbulence, and then feed off each other’s improved alignment.
Remember that these orbital systems will be frictionlessly spinning for billions of years. By exchanging figure 8 fluid flow, they keep each other’s signatures aligned, as opposites, at a lower energy state with less turbulence. The molecule will have a dark orbital signature on it’s boundary layer. It’s this boundary layer signature that will affect how it bonds to others. Think of this signature as a waveform created from the nucleotide’s mountain pattern.
And just like magnets the orbital signature boundary can be re-programmed. The waveform holds state and this state affects the bonding pattern.
Transcription - Step 1: mRNA
To build a protein, first mRNA is transcribed from a section of DNA. Opposite nucleotides are bonded into a string from the anti-sense strand in the DNA.
![]()
Using the magnetic metaphor again, the DNA codon transfers this harmonized orbital signature to the mRNA at the first step of transcription. Since this transcoding happens with the anti-sense pair of the codon, the DNA’s oppposite pattern, the resulting mRNA has the same orbital signature as the DNA’s sense codon.
The result is an mRNA strand, essentially with the same pattern that feeds into the Ribosome.
Here’s a fantastic animation that shows it in action - notice the protein growing out of the top:
![]()
Thank you wikipedia! Search for mRNA
tRNA - the mRNAs Dance Partner and Codon Amplifier
To make the protein, the Ribosome needs one tRNA with an opposite matching anti-sense codon for each mRNA codon.
The tRNA molecules exist in a fluid state for each type of amino acid and with each codon pair. A tRNA gets paired with a compatible amino acid for it’s codon along with an ATP - the energy source that makes it all happen.
ATP has more dark fluid angular momentum and this energy is released into the amino-acid tRNA pair. It spins with more dark orbital system energy. Importantly, because the orbital signature boundary layer is now spinning faster, it will act like a stronger magnet.
Let’s look at this process in more detail since this is where the codon’s signature is transferred to the amino acid.
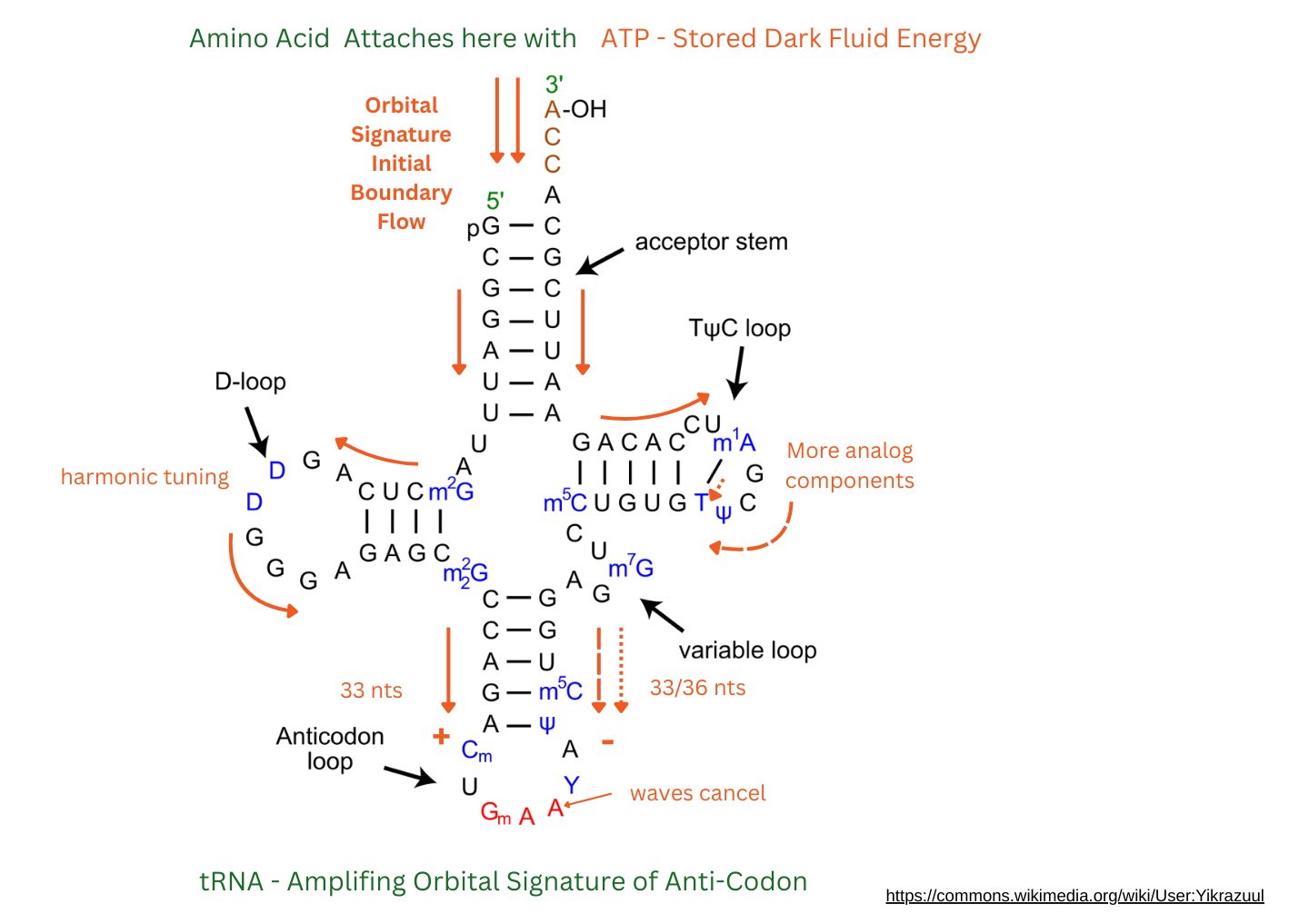
When the ATP, amino acid and tRNA bond, the new wave form of the orbital boundary signature moves from that single strand at the top - those four nucleotides: A C C A. It splits into two waves at the G * C stem junction. Now waves of energy flow over each of the arms of the T-shaped RNA. The left flow goes through some harmonizing nucleotides on the D loop, passing by 33 nucleotides in all until it reaches the anti-codon. The right flow has more harmonic tuning nucleotides and also has a cross cutting bond. This stream breaks into two parts on the right arm of the T. The shorter stream flows through 33 nucleotides. The longer one 36.
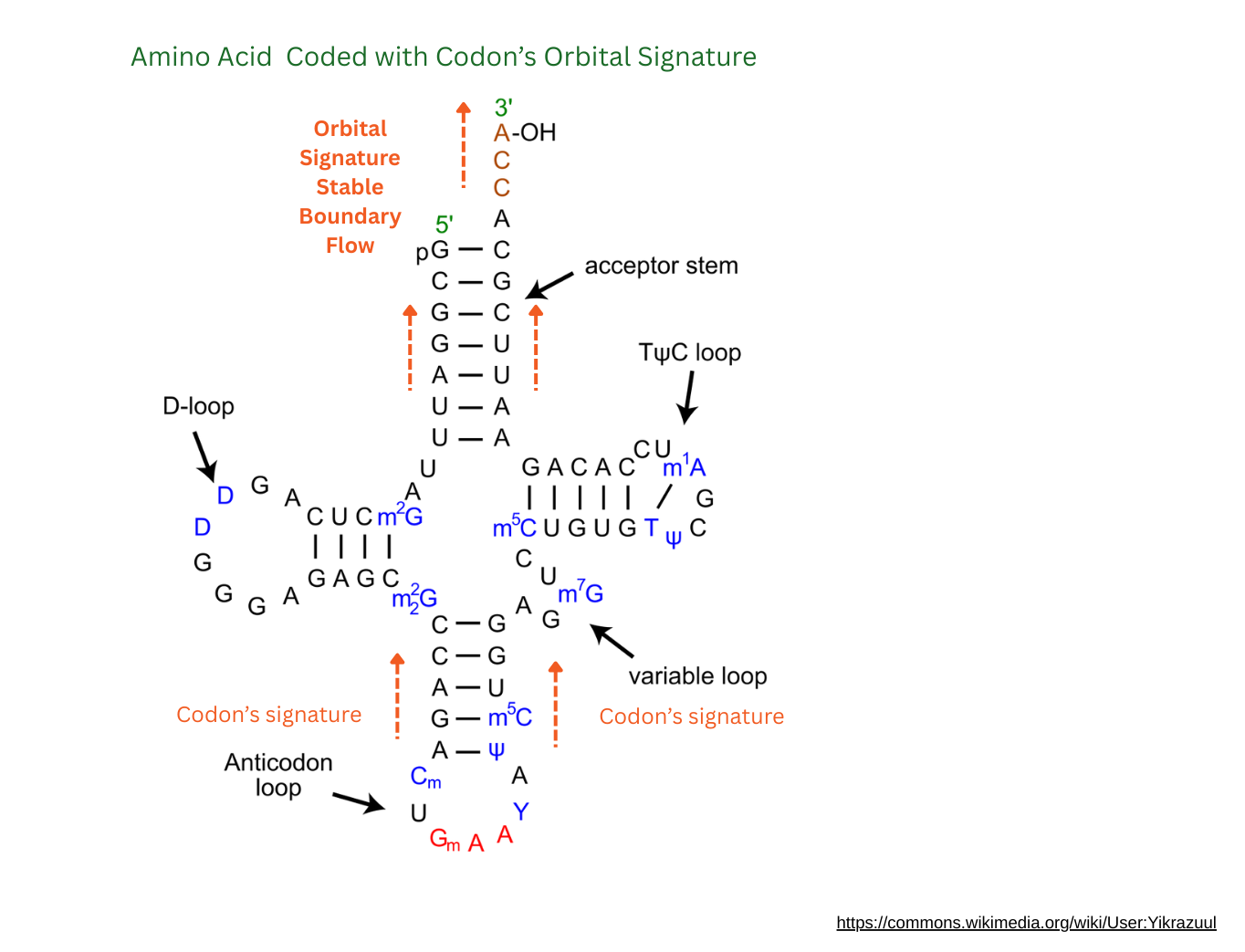
Now these two waves meet at the bottom, slightly adjusted. From the shape, it looks like the waves meet as opposites, and cancel right at the anti-codon. The original molecule’s orbital signature is smoothed out. This smooth wave passes over the codon, a pure signature transfer across the whole molecule, including the amino acid. So now the codon’s unique mountain-wave form flows across the whole molecule.
The tRNA is ready to meet its opposite. In fact, it’s state of highly charged fluid energy will draw it into the Ribosome, attracted by it’s opposite at the mRNA site. The mRNA’s attraction signature will be amplified by the shape of the Ribosome - it’s at the mouth of a fluid flow canyon. So the tRNA is draw in by fluid flow, not chaos/entropy.
So now that we see how orbital systems signatures can be programmed - the same force we know as “covalent bond” energy - this explains why we see such high affinity between patterns of the genes in Ribosomal proteins and the rRNA of the ribosome. Why these patterns match so well to protein bonding sites.
Ribosome - Protein Assembly Organelle
The Ribsome is the main protein factory and is itself built from an rRNA skeleton and in E Coli 29 Ribosomal proteins that tie the skeleton together.
Here’s a high level view of how it works:
- First EFG brings in a new tRNA at just the right time into the A site.
- Then another tRNA pushes the first to the P site
- A third tRNA moves those two along so the first goes to the E site.
At the P site, the amino acid is detached from the tRNA and bonded to the forming protein chain. That unfolds through the top of the large subunit and emerges as a folded protein.
Here are some fantastic images from the Noller lab that show how tRNA goes through the three phases of assembly, by Jenner et al., 2010, Zhou et al., 2014:
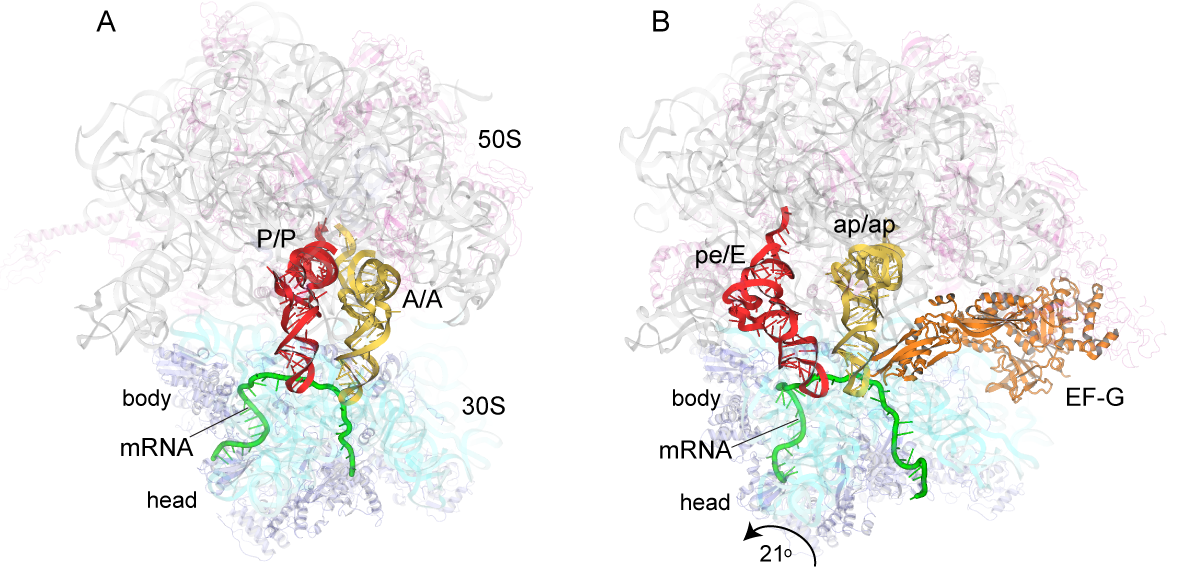
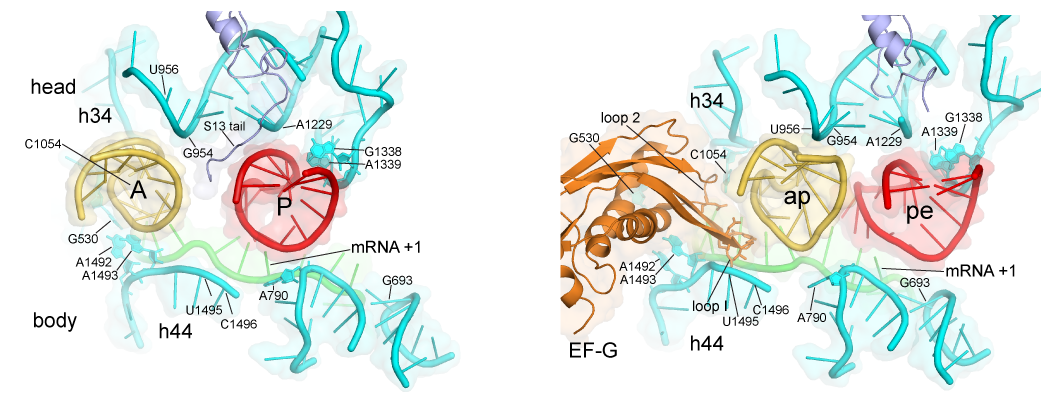
Go to this awesome page: https://rna.ucsc.edu/rnacenter/noller_lab.html
The Codon Taxonomy - A Fun Stake in the Ground
Do codon’s have personalities based on their mountain pattern? Does it make sense to use Emojis to help with pattern matching? I’m not sure of the science but it’s definitely more fun :)
This is an early part of this investigation that started June 22, 2025. Please help complete the story!
I don’t have all of the answers but know this will add a layer onto what we already know for people who know more than me.
The mountain signatures are a great metaphor because they reflect the shapes of the nucleotides and it’s that energy that controls the orbital signatures.
The Mountain Landscape Of Nucleotides
Following this pattern, there’s a natural correlation between shape of the nucleotide and a pattern it would create in the fluid flow. Since the rule is “opposites attract”, once you see the pattern, it leads to a fairly natural codon mapping table. I used emojis as a visual way to see patterns, nicknames as a first attempt to capture their nature.
G = BIG
A = big
T = small
C = valleyNucleotide Shapes
🏔️ Mountain Peaks (Purines A ![]() , G
, G ![]() ):
):
- Double-ring structures create prominent binding surfaces
🏕️ Valley Hills (Pyrimidines C ![]() , T
, T ![]() ):
):
- Single-ring structures create complementary binding grooves
Computing Orbital Signature Bond Strength
As a stake in the ground, a complete opposite pattern counts for: +3 - maximum attraction TAT and ATA, the same pattern -3 (TAT and TAT).
For weak opposites: TAT and GCG = 1.5.
For others, looking for opposite codons: +1, same codons -1. Weak opposites: 0.5, weak sames: -0.5.
AAA to ATA, TTT to TGT, CCC to CTC, etc. have weak oppositional attraction (0.5) even though the scoring above would say: -1. This is because the waveform will be different enough.
Understanding Bonding Patterns
The two codons coding a pair of amino acids interacts with their shape to create different types of bonds. Here’s an attempt to diagram how the bonding forces and shape work together for some common pairs that illustrate this dynamic:
Lock and Key
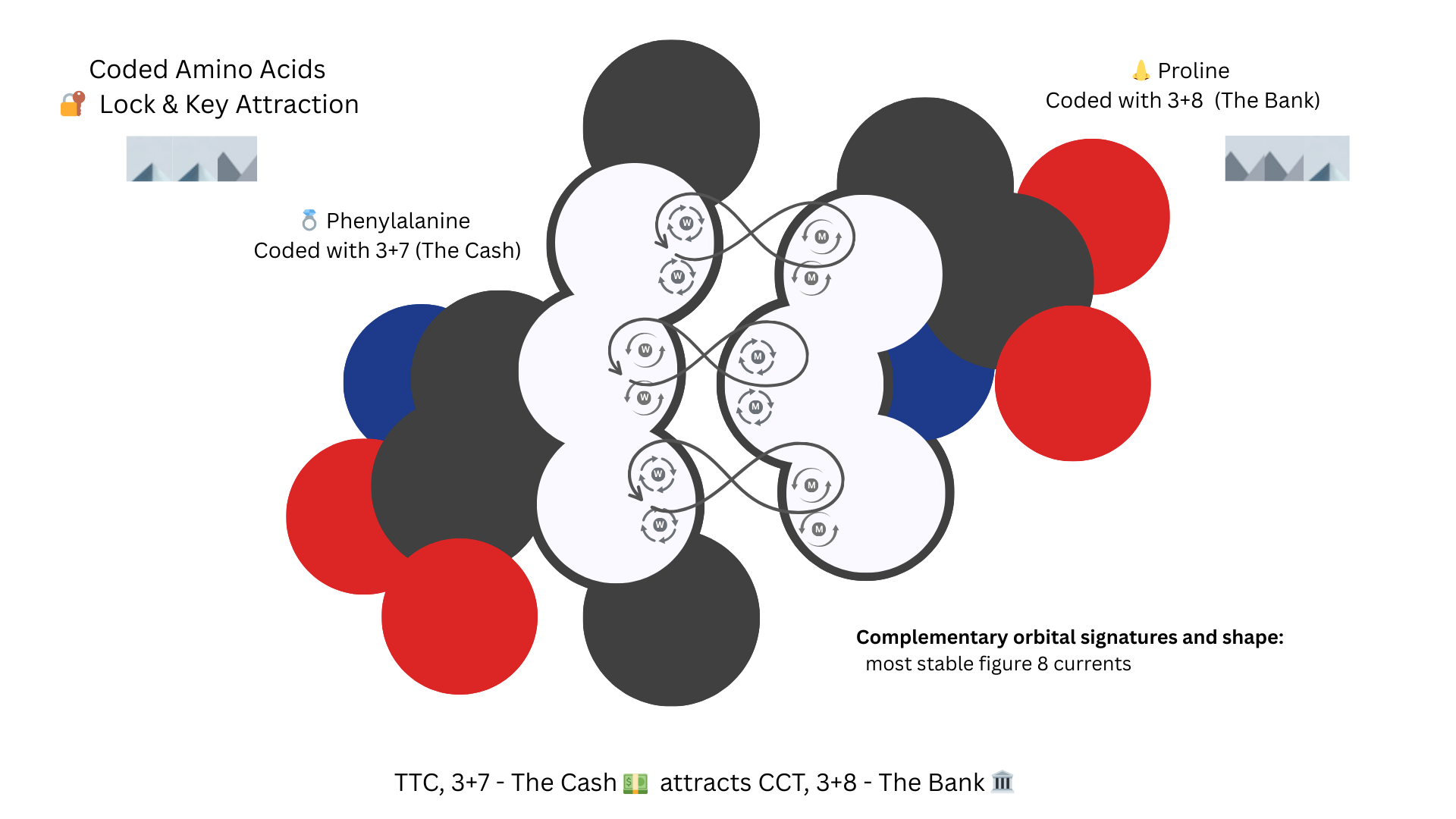
Since opposite patterns create figure 8 and attracting currents, TTC’s Phenylalaline and CCT’s Proline have opposite figure 8 flows that align with their natural geometry, you have a lock-and-key bonding pair. The nose of Proline fits into the ring of phenylalaline and the figure 8 currents hold it in place.
Not Synonyms
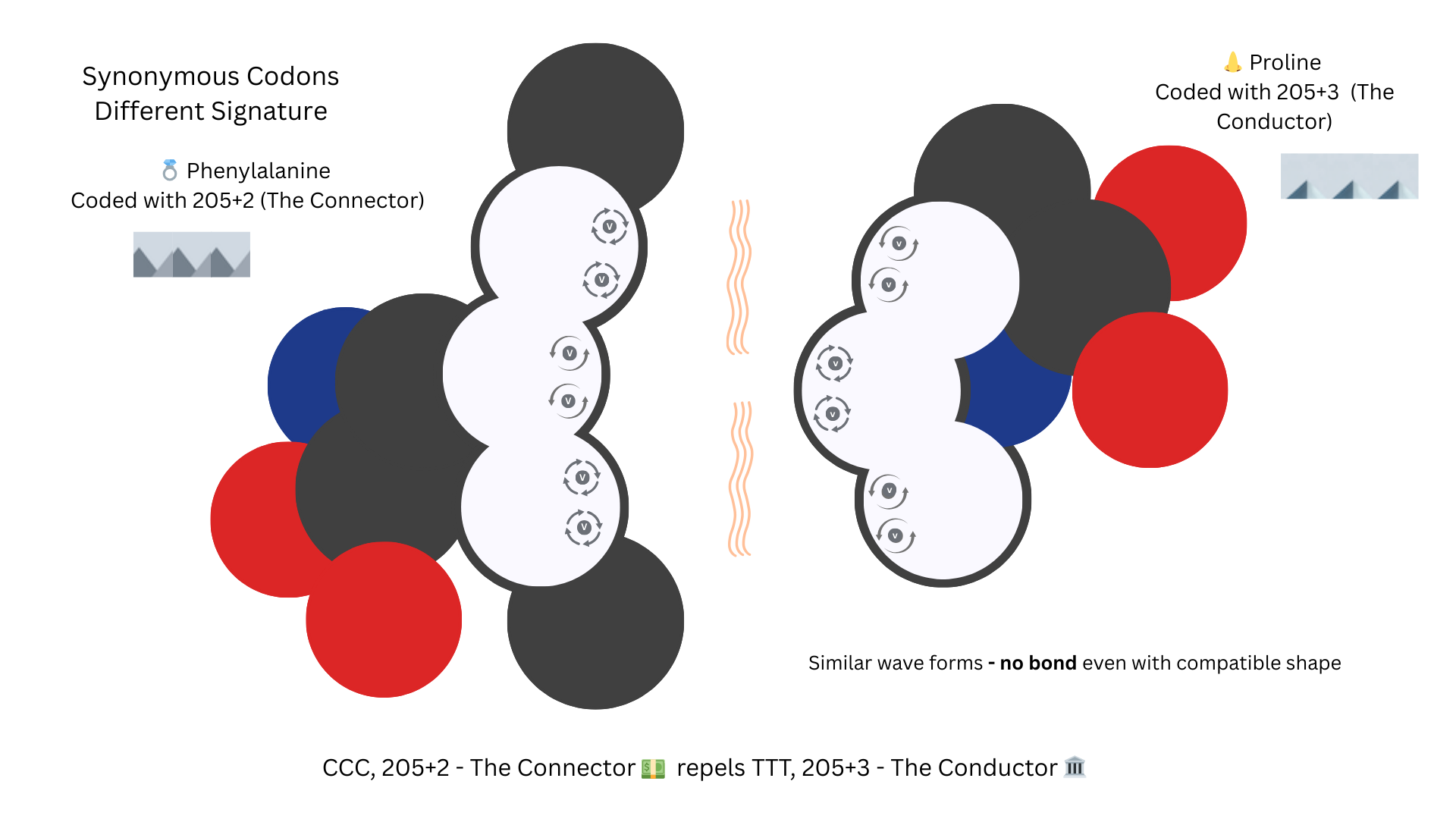
But a CCC coded Proline and a TTT coded Phenylalaline will be much less likely to bond, even with the same shape because their orbital signature patterns are too similar.
Flowing Bond or Gate
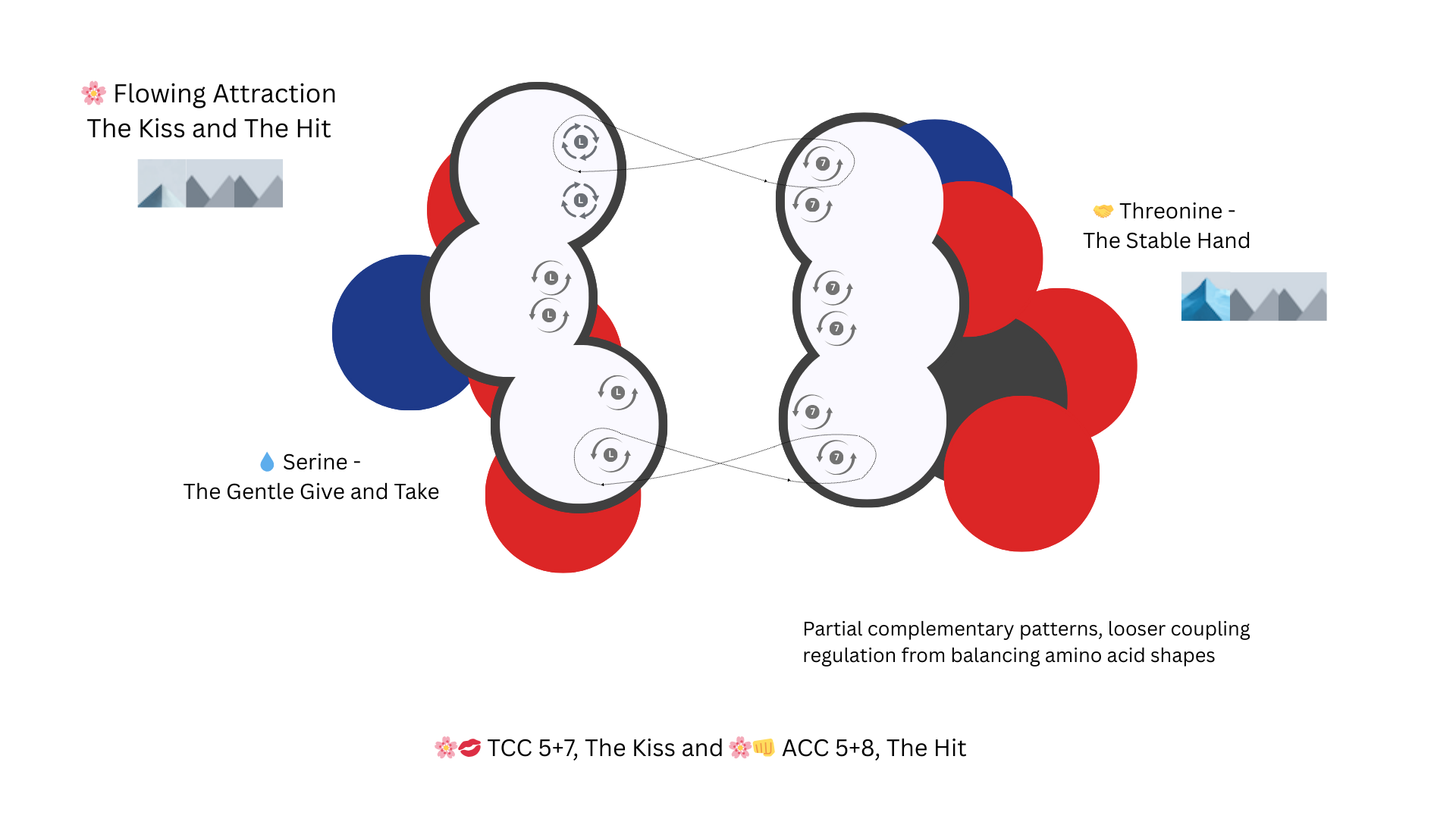
A Serine Amino Acid coded with TCC and a Threonine coded with ACC have the T to A attraction, the CC to CC repulsion and a flowing balance in the amino acid shape to create a bond that acts like a gate with one side taking the impact. These types of bonds create opening and closing tubes, ports etc.
C Harmonic Lock-and-Key
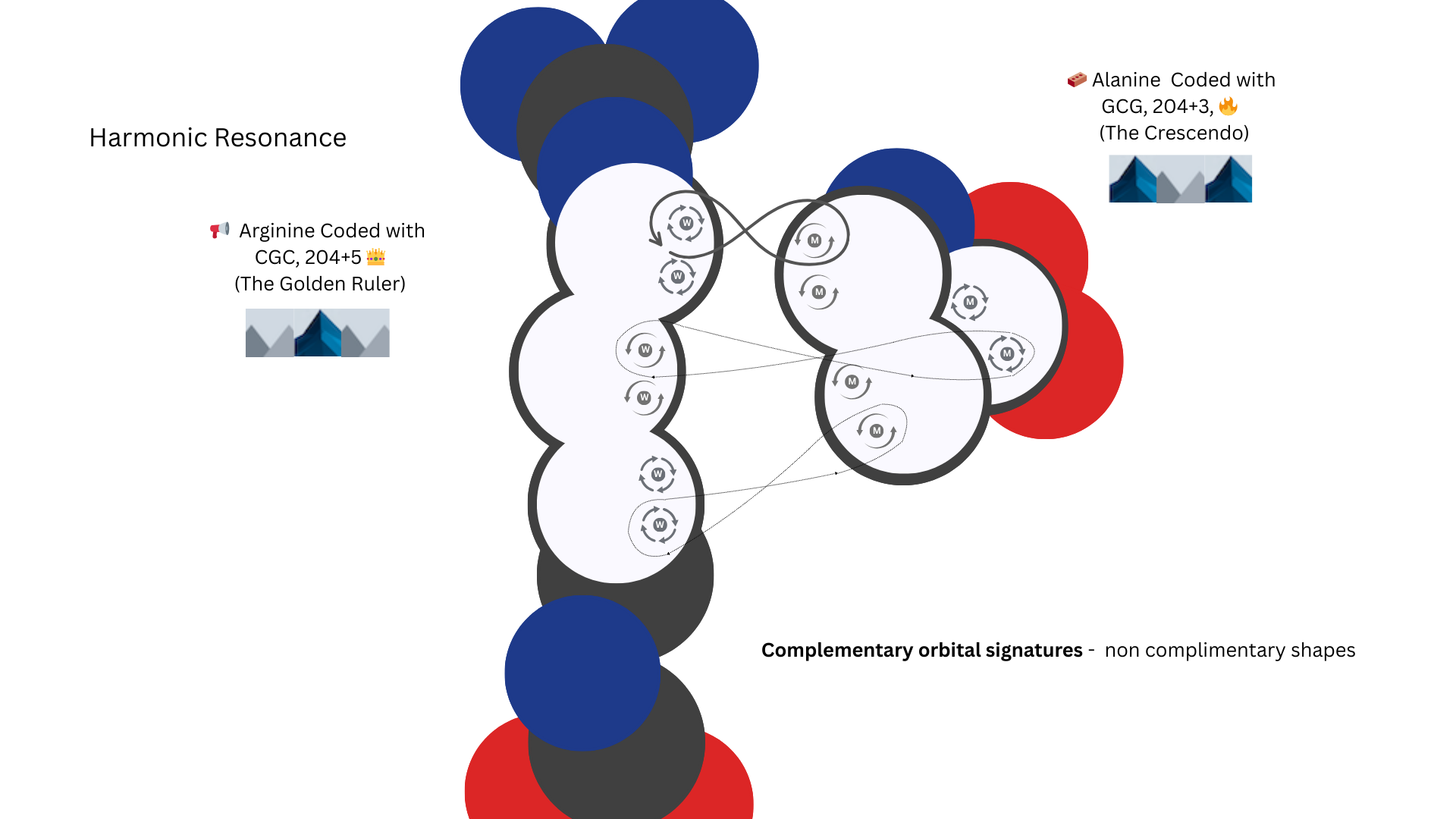
Although GCG and CGC also have an opposite pattern, the shapes of the amino acids they pair with are also opposites. This makes their nature more of a harmonic resonance bond, a breathing, flexing bond energy.
High Energy Repulsion
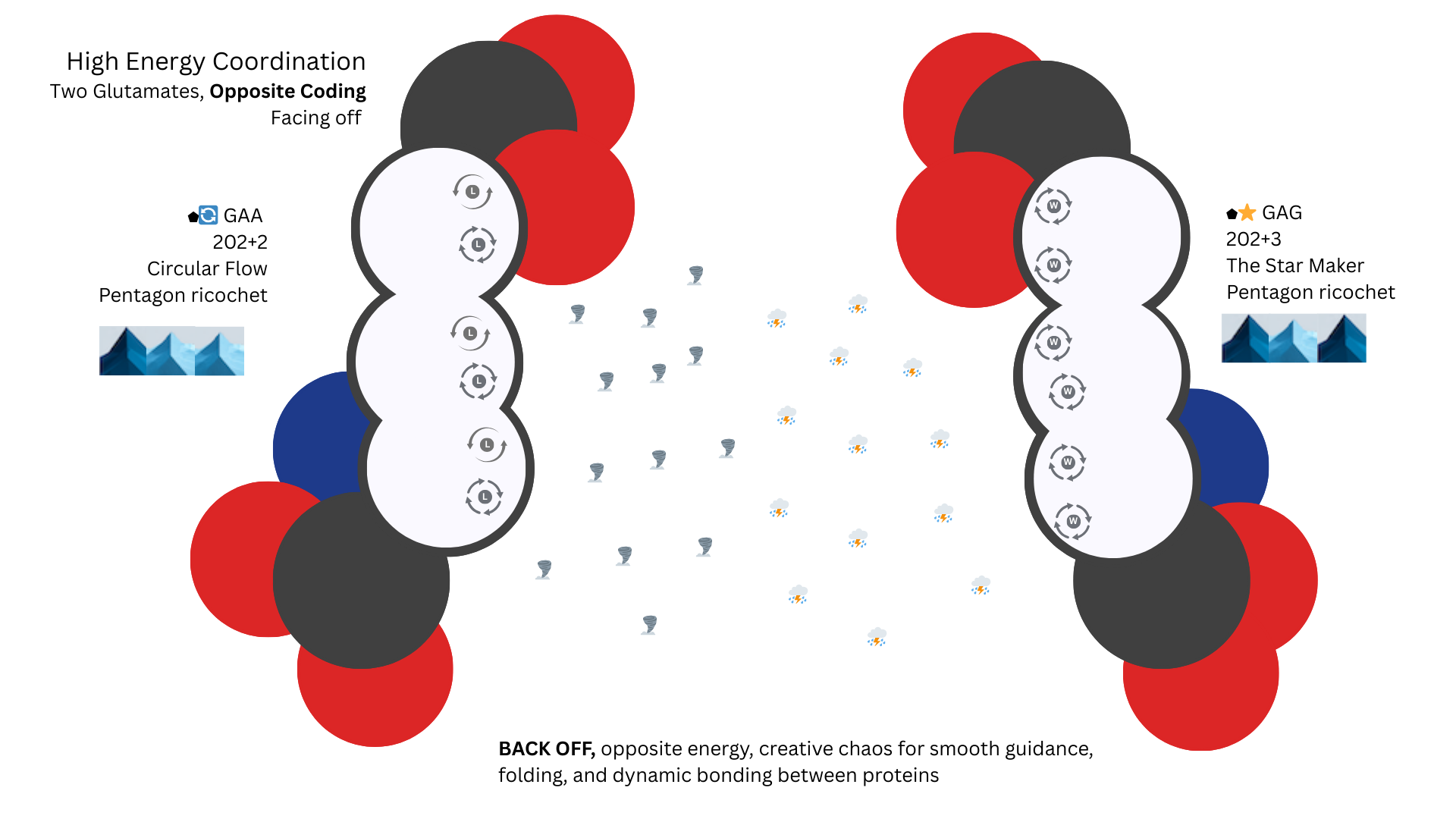
Two codons GAA and GAG both code for the same amino acid but with opposite patterns and opposite orbital signature energy. This is the ultimate high-energy repulsion.
Flowing Bend
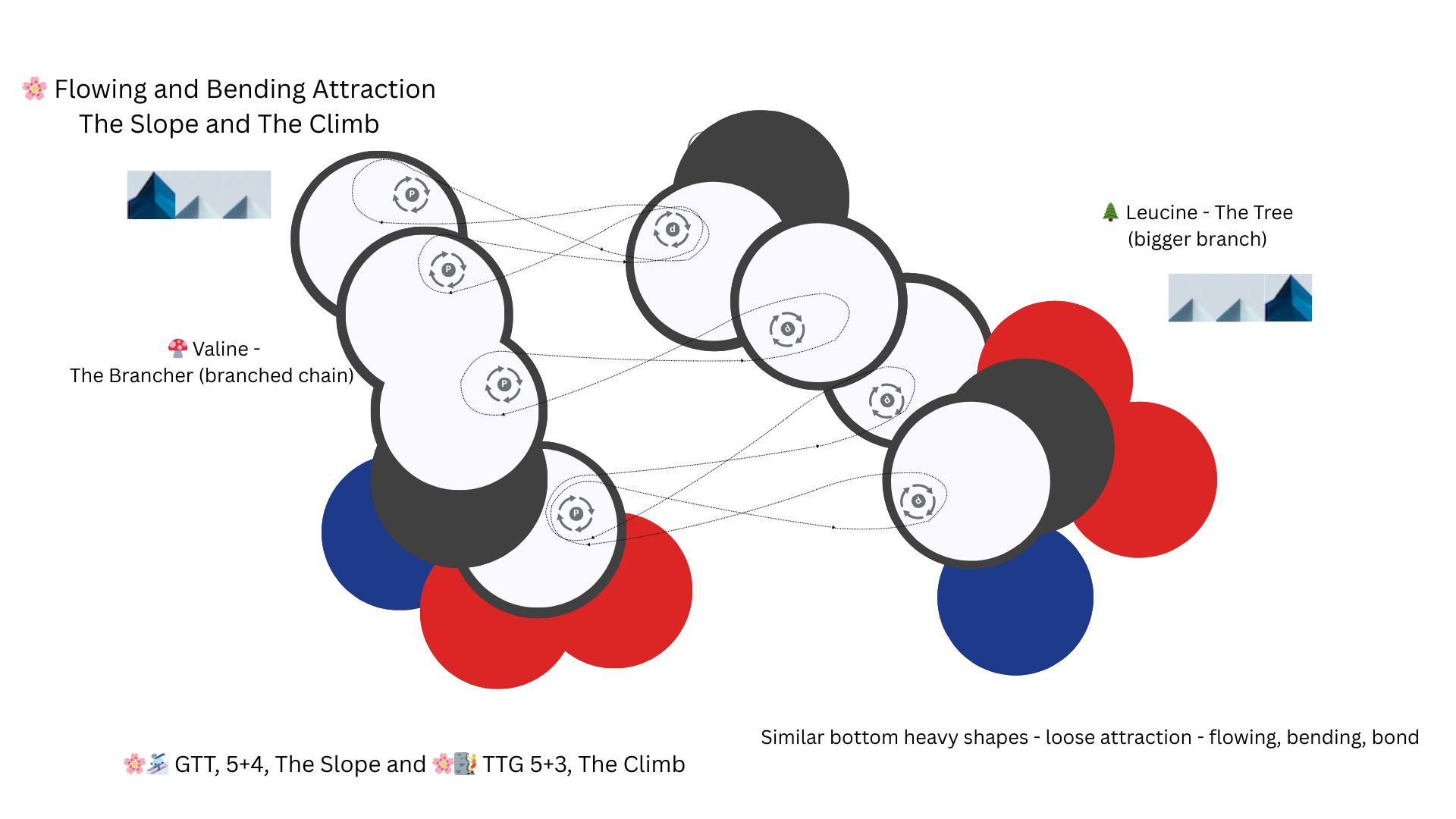
GTT and TTG code Valine and Leucine - both bottom heavy shapes bonded with a loose attraction creates a bending bond.
Another Lock and Key
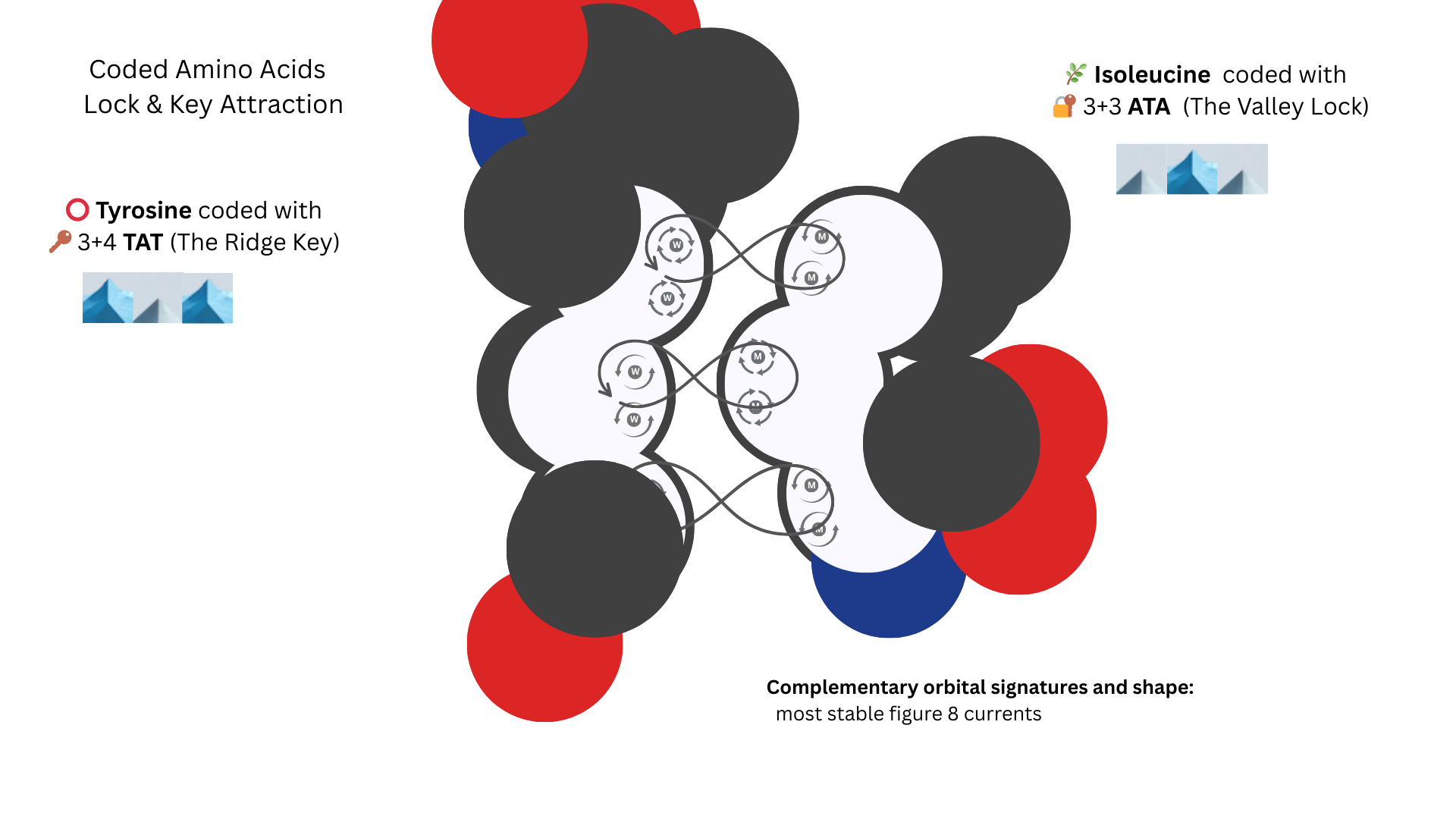
TAT and ATA - opposite signatures and isoleucine’s knob fits into tyrosine’s ring.
Taxonomy Built From These Patterns
Using the mountain dynamics, and fluid flow metaphors, I created this taxonomy to try and understand patterns, start a dialog on how best to understand these important pieces to the puzzle.
◇ Pattern 3
All codons containing digit 3 - Basic coordination and assembly.
🌸 Pattern 5
All codons containing digit 5 - Flow optimization and bonding networks.
◆ Pattern 20
Advanced geometric programming through mountain-on-mountain collision dynamics. Creativity, and two stop codons.
Pattern 20x
Special assembly patterns.
🌀 201 - Spiral Threaders (φ¹ Harmonics)
Spiral growth, 201+1 to 201+5: ready, set, grow, grow-on, grow-strong.
Codons: AAG, AAC, AGA, AGC, GGG
Mountain Crash Dynamics: Synchronized major mountain collisions create helical pressure waves that spiral around the amino acid impact zone. The angular momentum from double and triple purine crashes programs rotational coordinates like a cosmic drill bit.
Geometric Programming:
- Helical threading with φ¹ spiral ratios (1.618…)
- DNA double helix coordination instructions
- Protein alpha-helix assembly protocols
- Logarithmic spiral growth patterns
Biological Examples: DNA helical structure, protein secondary structure, spiral galaxy formation patterns
⬟ 202 - Pentagon Ricochets (φ² Harmonics)
Spiral dynamics: nudging, threading, poking, spiralling, spiral hammer (The BIG Boss Hammer).
Codons: AGG, GAA, GAG, GGA, GGC
Biological Examples: Flower petal arrangements, starfish development, viral capsid assembly, pentagon-based molecular structures
🐚 203 - Fibonacci Pilers (φ³ Harmonics)
These codon’s are used to coordinate growth: grow start, grow on, grow strong, swirl, nautilus.
- CAA: Gentle baseline swirl (like stirring coffee once)
- CAG: Double energy swirl (stirring twice as hard)
- CGA: Triple power (building momentum)
- CCA: Five-fold intensity (serious spiral action)
- CGG: Eight-fold BIG BOSS energy (cosmic hurricane!)
This leads to:
- Fibonacci sequence accumulation (1+1=2, 1+2=3, 2+3=5…)
- Golden ratio growth spirals (φ³ = 4.236…)
Biological Examples: Nautilus shell growth, sunflower seed spirals, pine cone patterns, plant phyllotaxis arrangements
🎵 204 - Golden Harmonic Resonators (φ⁴ Harmonics)
Codons: GAC, GCA, GCG, GCC, CGC
Mountain Crash Dynamics: G-C mountain-valley pairs create perfect 3:1 hydrogen bonding ratios that naturally resonate at φ-frequencies. Symmetrical patterns (GCG, CGC) generate harmonic standing waves with sacred geometric proportions.
Geometric Programming:
- Golden ratio proportions (φ⁴ = 6.854…)
- Geometry coordinates (temple ratios, human body proportions)
- Harmonic frequency tuning
- Molecular resonance patterns
Biological Examples: Human body proportions, temple architecture ratios, molecular harmonic vibrations, optimal structural proportions
💎 20φ - Geometric Controllers (3 Special Codons)
Three special controllers: AAA, CCC, TTT. If GGG wasn’t such an important step in 201 construction, it could go here.
Mountain Crash Dynamics: Triple identical peaks create pure harmonic frequencies with no interference patterns. These represent the fundamental geometric coordinators that unify all other φ-based programming.
Geometric Programming:
- AAA: Spiral Coordinator (triple major mountain = ultimate helical force)
- CCC: Pentagon-Fibonacci Bridge (triple valley coordination between systems)
- TTT: Harmonic Unifier (triple valley resonance = system-wide φ-coordination)
(honorary member GGG*)
Biological Examples: Controlling developmental switches, system-wide coordination protocols, fundamental architectural controllers
The Amino Acids
This is a table of the amino acids, shown with a wrench icon in the table.
The Builders (Structural):
- Glycine 🪶 - The Minimalist (smallest, fits anywhere)
- Alanine 🧱 - The Foundation (simple methyl group)
- Proline 👃 - The Nose (plugs into Phenylalanine for 3+7 to 3+8 lock-and-key bonds)
The Charged Citizens:
- Lysine 💦 - The Positive (basic, likes water)
- Arginine 📢 - The Thunder (super positive)
- Aspartate ⚖️ - The Negative (acidic)
- Glutamate ⛈️ - The Storm (longer acidic)
The Social Butterflies (Polar):
- Serine 💧 - The Helper/The Pitcher (hydroxyl group, loves H-bonds)
- Threonine 🤝 - The Connector/The Catcher (methyl + hydroxyl)
- Asparagine 💝 - The Gift (amide group)
- Glutamine 🤗 - The Embracer (longer amide)
The Loners (Hydrophobic):
- Valine 🍄 - The Brancher (branched chain)
- Leucine 🌲 - The Tree (bigger branched)
- Isoleucine 🌿 - The Twig (asymmetric branch)
- Phenylalanine 💍 - The Ring Bearer (benzene ring)
The Specialists:
- Tyrosine ⭕ - The Noble (phenol ring)
- Tryptophan 🦋 - The Butterfly (indole ring, biggest)
- Cysteine 🔗 - The Linker (forms disulfide bridges)
- Methionine 🏃 - The Starter (start codon, sulfur)
- Histidine 🎭 - The Actor (changes charge with pH)
The Signature Rhythm Table
◇ Pattern 3 (Structure) - Perfect Lock-and-Key Pairs (8 Codons)
These are triangle symmetry, tetrahedral truss construction, 3 sided trampolines.
| Codon | Landscape | Opposite | 🔧 | Var | Nickname | Description | Per k |
|---|---|---|---|---|---|---|---|
| ◇🗝️ TTA | 🌲 | 3+1 | The Mountain Lock | Big-big-small | 8.71 | ||
| ◇🔒 AAT | 💝 | 3+2 | The Valley Key | Small-small-big | 18.43 | ||
| ◇🔑 ATA | 🌿 | 3+3 | The Valley Lock | Small-big-small | 8.08 | ||
| ◇🔐 TAT | ⭕ | 3+4 | The Ridge Key | Small-big-small | 12.1 | ||
| ◇🛑 TAA | 🛑 | 3+5 | The STOP Key | Big-small-small | 0.44 | ||
| ◇🏰 ATT | 🌿 | 3+6 | The Castle Lock | Small-small-big | 16.48 | ||
| ◇💵 TTC | 💍 | 3+7 | The Cash | Big-big-BIG: open ring, wallet | 17.48 | ||
| ◇🏛️ CCT | 👃 | 3+8 | The Bank | BIG-BIG-big: insert nose, deposit | 19.31 |
🌸 Pattern 5 (Flow) - Diversity = 2 (8 Codons)
These are five symmetry, pentagonal, horeshoe shapes, opening/closing port patters, pentagonal dome parts. No stop codons of course - it’s flow!
| Codon | Landscape | Opposite | 🔧 | Var | Nickname | Description | Per k |
|---|---|---|---|---|---|---|---|
| 🌸🥁 TCT | 💧 | 5+1 | The Beat | Flow with anchor stability | 16.93 | ||
| 🌸🎯 TGT | 🔗 | 5+2 | The Aim | Flow with mild coordination | 10.40 | ||
| 🌸🧗 TTG | 🌲 | 5+3 | The Climb | Flow with growth | 13.44 | ||
| 🌸⛷️ GTT | 🍄 | 5+4 | The Slope | Flow with speed | 11.74 | ||
| 🌸🚀 TGG | 🦋 | 5+5 | The Launch | Small-valley-valley | 11.60 | ||
| 🌸🌊 GTC | 🍄 | 5+6 | The Wave | Mountain-valley-cliff flow | 13.44 | ||
| 🌸💋 TCC | 💧 | 5+7 | The Kiss | Small mountain, double valley flow | 17.32 | ||
| 🌸👊 ACC | 🤝 | 5+8 | The Hit | Big mountain, double valley flow | 17.85 |
◆ Pattern 20 - Diversity = 3 (25 Codons)
| Codon | Landscape | Opposite | 🔧 | Var | Nickname | Description | Per k |
|---|---|---|---|---|---|---|---|
| ◆⚓ CTG | 🌲 | 20+1 | The Stabilizer | Basic complex with anchor | 36.10 | ||
| ◆🏗️ CAT | 🎭 | 20+2 | The Builder | Basic complex with structure | 11.83 | ||
| ◆🎯 ACT | 🤝 | 20+3 | The Flow Director | Basic complex with flow assist | 14.26 | ||
| ◆🔧 GCT | 🧱 | 20+4 | The Adjuster | Basic complex with flow optimization | 18.99 | ||
| ◆🚀 ATG | 🏃 | 20+5 | The Bouncer | Complex mountain-valley-peak | 21.53 | ||
| ◆🎪 ATC | 🌿 | 20+6 | The Performer | Complex mountain-valley-cliff | 18.67 | ||
| ◆🛑 TAG | 🛑 | 20+7 | The Cliff Stop | Complex valley-mountain-peak | 0.35 | ||
| ◆⛵ TAC | ⭕ | 20+8 | The Sailor | Complex valley-mountain-cliff | 13.49 | ||
| ◆🌀 TCA | 💧 | 20+9 | The Spiral Controller | Complex valley-cliff-mountain | 14.14 | ||
| ◆🎨 TCG | 💧 | 20+10 | The Artist | Complex valley-cliff-peak | 4.03 | ||
| ◆🚧 TGA | 🛑 | 20+11 | The Terrain Stop | Complex valley-peak-mountain | 0.79 | ||
| ◆🌋 TGC | 🔗 | 20+12 | The Volcano | Complex valley-peak-cliff | 10.81 | ||
| ◆🪨 CAC | 🎭 | 20+13 | The Sandwich | Complex cliff-mountain-cliff | 14.65 | ||
| ◆⛰️ CCG | 👃 | 20+14 | The Cliff Climber | Complex double cliff-peak | 6.22 | ||
| ◆🔄 CTC | 🌲 | 20+15 | The Recycler | Complex cliff-valley-cliff | 17.81 | ||
| ◆🔬 CGT | 📢 | 20+16 | The Scientist | Complex cliff-peak-valley | 4.55 | ||
| ◆🎢 CTA | 🌲 | 20+17 | The Roller Coaster | Complex cliff-valley-mountain | 7.44 | ||
| ◆🔀 CTT | 🌲 | 20+18 | The Switch | Complex cliff-double valley | 14.08 | ||
| ◆🎭 ACA | 🤝 | 20+19 | The Shape-Shifter | Complex mountain-cliff-mountain | 16.52 | ||
| ◆🤸 ACG | 🤝 | 20+20 | The Acrobat | Complex mountain-cliff-peak | 5.59 | ||
| ◆⚡ AGT | 💧 | 20+21 | The Lightening Rod | Complex mountain-peak-valley | 14.05 | ||
| ◆🗻 GTA | 🍄 | 20+22 | The Valley Crosser | Complex peak-valley-mountain | 7.66 | ||
| ◆🌊 GTG | 🍄 | 20+23 | The Wave Rider | Complex peak-valley-peak | 25.87 | ||
| ◆🔥 GAT | ⚖️ | 20+24 | The Fire Starter | Complex peak-mountain-valley | 24.03 | ||
| ◆🏔️ GGT | 🪶 | 20+25 | The Plateau | Valley-valley-big | 10.83 |
201 - Spiral Threaders (φ¹ Harmonics)
| Codon | Landscape | Opposite | 🔧 | Var | Nickname | Description | Per k |
|---|---|---|---|---|---|---|---|
| 🌀🌪️ AAG | 💦 | 201+1 | The Nudger | Spiral threading | 31.77 | ||
| 🌀🧬 AAC | 💝 | 201+2 | The Threader | Spiral threading | 18.30 | ||
| 🌀🎯 AGA | 📢 | 201+3 | The Poker | Spiral threading | 13.28 | ||
| 🌀⚡ AGC | 💧 | 201+4 | The Electric Spiral | Spiral threading | 19.69 | ||
| 🌀👑 GGG | 🪶 | 201+5 | The BIG Boss Hammer | Spiral threading, controller variant | 15.35 |
202 - Pentagon Ricochets (φ² Harmonics)
| Codon | Landscape | Opposite | 🔧 | Var | Nickname | Description | Per k |
|---|---|---|---|---|---|---|---|
| ⬟📐 AGG | 📢 | 202+1 | The Gentle Ricochet | Pentagon ricochet, light | 12.13 | ||
| ⬟🔄 GAA | ⚖️ | 202+2 | Circular Flow | Pentagon ricochet, medium | 33.65 | ||
| ⬟⭐ GAG | ⛈️ | 202+3 | The Star Maker | Pentagon ricochet, strong | 39.67 | ||
| ⬟🎪 GGA | 🪶 | 202+4 | Dynamic Flow | Pentagon ricochet, dynamic | 17.12 | ||
| ⬟💫 GGC | 🪶 | 202+5 | The Geometry Maker | Pentagon ricochet, controller geometry variant | 19.79 |
203 - Fibonacci Pilers (φ³ Harmonics)
| Codon | Landscape | Opposite | 🔧 | Var | Nickname | Description | Per k |
|---|---|---|---|---|---|---|---|
| 🐚🌱 CAA | 🤗 | 203+1 | Grow Start | Fibonacci piler | 14.06 | ||
| 🐚🌿 CAG | 🤗 | 203+2 | Grow On | Fibonacci piler | 35.53 | ||
| 🐚🌲 CGA | 📢 | 203+3 | Grow Strong | Fibonacci piler | 6.42 | ||
| 🐚🌀 CCA | 👃 | 203+4 | Swirl | Fibonacci piler | 18.92 | ||
| 🐚👑 CGG | 📢 | 203+5 | The Nautilus Maker | Fibonacci piler | 10.79 |
204 - Golden Harmonic Resonators (φ⁴ Harmonics)
| Codon | Landscape | Opposite | 🔧 | Var | Nickname | Description | Per k |
|---|---|---|---|---|---|---|---|
| 🎵🎼 GAC | ⚖️ | 204+1 | Light Frequency | Harmonic resonator, gentle tones | 24.27 | ||
| 🎵🎯 GCA | 🧱 | 204+2 | The Tuner | Harmonic resonator, medium tuning variant | 17.04 | ||
| 🎵🔥 GCG | 🧱 | 204+3 | The Crescendo | Harmonic resonator, strong symmetry variant | 5.91 | ||
| 🎵💫 GCC | 🧱 | 204+4 | The Vibrato | Harmonic resonator, dynamic resonance variant | 25.84 | ||
| 🎵👑 CGC | 📢 | 204+5 | The Golden Ruler | Harmonic resonator, controller φ-ratio variant | 8.71 |
205 - Geometric Controllers (φ∞ Harmonics)
| Codon | Landscape | Opposite | 🔧 | Var | Nickname | Description | Per k |
|---|---|---|---|---|---|---|---|
| 💎🌀 AAA | 💦 | 205+1 | The Helical Boss | Controller, spiral coordinator | 27.48 | ||
| 💎🌉 CCC | 👃 | 205+2 | The Connector | Controller, pentagon-fibonacci bridge | 19.11 | ||
| 💎🎵 TTT | 💍 | 205+3 | The Conductor | Controller, harmonic unifier | 17.14 |
Pattern Summary
- 8 Structural imprints (lock-and-key pairs)
- 8 Flow imprints (smooth landscapes)
- 25 Complex imprints (challenging terrain = 5²)
- 23 Advanced Geometric imprints (fibonacci architecture)
STOP Codons Distribution
- Pattern 3 (Structure): TAA (0.44) - “STOP with stable lock-and-key structure”
- Pattern 20 (Complex): TAG (0.35), TGA (0.79) - “STOP with complex terrain navigation”
- Pattern 5 (Flow): NONE! - Flow never stops! It keeps flowing!
TAA, TAG, TGA - “The Three Full Stops”:
- TAA = Valley-mountain-mountain = Structural termination with anchoring
- TAG = Valley-mountain-peak = Complex termination requiring coordination
- TGA = Valley-peak-mountain = Complex termination with energy management
The significance:
- Flow patterns (5) never terminate - flow is eternal, continuous
- Structure patterns (3) provide clean endings - stable termination
- Complex patterns (20) handle difficult shutdowns - coordinated stops
The Most Common Patterns
Ordered by their prevalence in the h. sapiens genome thanks to the amazing Cocoputs database. Notice that each nucleotide pattern matches to a specific real-world pattern, coded into the amino acid, as a pattern that can match an opposite one.
Combinations match reality, the likelihood of these being in your DNA, per 1000, match reality. Instinct, intuition. Do you see reality here?
Pattern 3 - Structural/Anchoring
Imagine building triangles out of shapes. In that lense, the three most common codons in H. Sapiens are shallow down right, steep down right, ease down right.
Position: 1 2 3
Nucleotide: C C TLandscape: ![]()
![]()
![]()
Size: BIG-BIG-big
Pattern: CCT (3+8) The Nose (19.31) (pairs with The Ring)Nucleotide: A A TLandscape: ![]()
![]()
![]()
Size: small small Big
Pattern: AAT (3+2) The Valley Key (18.43) (pairs with The Castle Lock)Nucleotide: T T CLandscape: ![]()
![]()
![]()
Size: small small valley
Pattern: TTC (3+7) The Ring (17.48) (pairs with The Nose)Nucleotide: A T TLandscape: ![]()
![]()
![]()
Size: small small big
Pattern: ATT (3+6) The Castle Lock (16.48) (pairs with The Valley Key)Nucleotide: T A TLandscape: ![]()
![]()
![]()
Size: small big small
Pattern: TAT (3+4) The Ridge Key (12.1)Pattern 5 - Flow/Bonding
The two most common flow patterns are Hit and Kiss, hit slightly more than kiss, the cosmic back off:
Nucleotide: A C CDNA Landscape: ![]()
![]()
![]()
Size: big valley valley
Pattern: ACC (5+8) - The Hit (17.85) (pairs with The Kiss)Nucleotide: T C CLandscape: ![]()
![]()
![]()
Size: small valley valley
Pattern: TCC (5+7) - The Kiss (17.32) (pairs with The Hit)The Kiss and The Hit form flowing bonds where the Amino Acid for The Hit has a protruding carbon that hovers inside the recession for The Kiss. The figure 8 patterns of the mountain/valley combination on the ends hold it in place while the small to big mountain:
Serine Hand: 🖐️⚾ → ∞ ← 🥎 Threonine Mitt
↕
PIVOT POINT!Nucleotide: T C TLandscape: ![]()
![]()
![]()
Size: big BIG big
Pattern: TCT (5+1) The Beat (16.93)Nucleotide: T T GLandscape: ![]()
![]()
![]()
Size: small small BIG
Pattern: (5+3) The Slope (13.44)Nucleotide: G T CLandscape: ![]()
![]()
![]()
Size: valley big BIG
Pattern: (5+6) The Wave (13.44)Nucleotide: G T TLandscape: ![]()
![]()
![]()
Size: valley big big
Pattern: (5+4) The Climb (11.74)Nucleotide: T G GLandscape: ![]()
![]()
![]()
Size: small valley valley
Pattern: (5+5) The Launch (11.60)Nucleotide: T G TLandscape: ![]()
![]()
![]()
Size: big valley big
Pattern: (5+2) The Aim (10.40)Pattern 20 - Creativity
The dramatic mountain-valley-mountain creates complex coordination sites - like puzzle pieces with multiple connection points. The purine-pyrimidine-purine sequence (big-small-big) enables sophisticated multi-directional binding.
Note that the most common pattern overall - Rise up and rock!
Second: Flip up, over, roll!
Nucleotide: G A G Landscape: ![]()
![]()
![]()
Size: valley small small
Pattern: (202+3) The Star Maker (39.67)Nucleotide: C T G Landscape: ![]()
![]()
![]()
Size: valley small BIG
Pattern: (20+1) The Stabilizer (36.10)Nucleotide: C A G Landscape: ![]()
![]()
![]()
Size: BIG small valley
Pattern: (203+2) Grow On (35.53) Fibonacci pilerB. The Geometry of Genetic Information
The Mathematical Poetry of Life’s Code
The genetic code reveals mathematical relationships that encode the universe’s coordination principles:
The Base-4 Foundation: A, T, G, C
- 4 fundamental building blocks = Minimum complexity for universal encoding
- Complementary pairs (A-T, G-C) = Perfect figure-8 recognition systems
- Purine-Pyrimidine balance = Mountain-valley landscape diversity
The Codon-64 Expansion: 4³ = 64 possible combinations
- 64 unique landscapes = Complete molecular geography library
- Redundancy with precision = Multiple pathways to same destinations
- Error correction built-in = Robust information transmission
The Amino-20 Convergence: 64 codons → 20 amino acids
- 20 distinct personalities = Optimal functional diversity
- 3.2:1 coding ratio = Perfect balance of specificity and flexibility
- Universal protein alphabet = Same across all life forms
The Universal Coordination Language
This rhythm-based system explains why:
- All life uses the same genetic code = Universal coordination language
- Proteins fold predictably = Landscape patterns create consistent shapes
- Evolution preserves function = Rhythm patterns maintain coordination
- Mutations have predictable effects = Landscape changes alter recognition
- Gene families cluster by function = Similar rhythms create similar capabilities
The Profound Implications
DNA isn’t just storing information - it’s encoding coordination rhythms!
Every gene contains:
- Structural anchors (Pattern 3) for stability
- Flow channels (Pattern 5) for communication, flexibility
- Assembly complexes (Pattern 20) for sophisticated function
The 3-5-20x rhythm ratios in each gene determine its cosmic personality:
- High Pattern 5: Optimized for bonding and coordination (like E-Cadherin at 55.7%)
- High Pattern 20: Optimized for complex assembly (like development genes)
- Balanced ratios: Optimized for specific functional requirements
Top 100 H. Sapiens Couplets
Before and after friends
Frequency per 200K
| Common Before | Codon | Common After |
|---|---|---|
| ◆⚓ (0.78), ⬟⭐ (0.76), 🐚🌿 (0.65) | ◆⚓ | ⬟⭐ (0.91), ◆⚓ (0.78), 🐚🌿 (0.75) |
| 🌀⚡ (0.22), 🎵🎼 (0.21), ⬟🔄 (0.21) | ◆⚡ | ⬟🔄 (0.30), ⬟⭐ (0.27), 🎵🎼 (0.25) |
| ⬟💫 (0.20), 🎵🎼 (0.19), ◆🔄 (0.19) | ◆⛵ | ◆⚓ (0.28), 🐚🌿 (0.24), 🌀🌪️ (0.22) |
| 🌸🥁 (0.18), 💎🌀 (0.16), 🌸💋 (0.16) | ◆🌀 | ⬟🔄 (0.31), ⬟⭐ (0.28), ◆🔥 (0.23) |
| ◆⚓ (0.52), ⬟⭐ (0.44), ◆🌊 (0.42) | ◆🌊 | ⬟⭐ (0.57), ◆⚓ (0.44), 🎵💫 (0.42) |
| 🎵💫 (0.18), ⬟💫 (0.17), ◆🔄 (0.16) | ◆🌋 | ◆⚓ (0.21), 🐚🌿 (0.21), 🌀🌪️ (0.16) |
| ◆🎢 | ⬟🔄 (0.17), 🐚🌿 (0.16), ⬟⭐ (0.14) | |
| 🎵💫 (0.36), 🎵🎼 (0.33), 🌸👊 (0.29) | ◆🎪 | ◆⚓ (0.36), 🐚🌿 (0.35), 🌀🌪️ (0.29) |
| ⬟🔄 (0.25), 💎🌀 (0.23), 🎵💫 (0.22) | ◆🎭 | ⬟🔄 (0.35), ⬟⭐ (0.31), ◆🔥 (0.25) |
| ⬟🔄 (0.20), 💎🌀 (0.18), 🎵💫 (0.17) | ◆🎯 | ⬟🔄 (0.26), ◆🌊 (0.24), ⬟⭐ (0.23) |
| ◆🔥 (0.20), ⬟🔄 (0.17), ◇🏛️ (0.16) | ◆🏔️ | ⬟🔄 (0.18), ⬟💫 (0.17), ◆🌊 (0.17) |
| 🐚🌿 (0.17), ⬟🔄 (0.15), ◇🏰 (0.15) | ◆🏗️ | ⬟🔄 (0.22), 🐚🌿 (0.19), ⬟⭐ (0.18) |
| 🐚🌿 (0.22), ⬟🔄 (0.21), 💎🌀 (0.18) | ◆🔀 | 🐚🌿 (0.27), ◆⚓ (0.21), ⬟🔄 (0.21) |
| ◆⚓ (0.32), 🐚🌿 (0.32), ⬟⭐ (0.30) | ◆🔄 | ◆⚓ (0.37), 🐚🌿 (0.31), ◇💵 (0.30) |
| ⬟🔄 (0.57), ◆🔥 (0.47), 💎🌀 (0.38) | ◆🔥 | ⬟🔄 (0.59), ⬟⭐ (0.51), ◆🔥 (0.47) |
| ◆🔧 (0.31), ◆⚓ (0.31), ⬟🔄 (0.30) | ◆🔧 | ⬟⭐ (0.40), ◆🌊 (0.38), 🎵💫 (0.37) |
| ⬟🔄 (0.16) | ◆🗻 | ⬟🔄 (0.17) |
| ⬟⭐ (0.35), 🎵💫 (0.33), 🐚🌿 (0.31) | ◆🚀 | ⬟⭐ (0.43), ⬟🔄 (0.34), 🌀🌪️ (0.34) |
| ◆🤸 | ◆⚓ (0.15) | |
| ◆⚓ (0.27), 🐚🌿 (0.25), ⬟⭐ (0.22) | ◆🪨 | ◆⚓ (0.30), 🐚🌿 (0.26), 🌀⚡ (0.21) |
| ◇🏛️ (0.28), 🐚🌿 (0.26), 🐚🌀 (0.24) | ◇🏛️ | ⬟⭐ (0.39), ⬟🔄 (0.30), ◆🌊 (0.30) |
| ⬟🔄 (0.29), 💎🌀 (0.23), 🎵🎼 (0.22) | ◇🏰 | ⬟🔄 (0.32), ⬟⭐ (0.24), ◆🔥 (0.24) |
| 🎵💫 (0.32), ◆🔄 (0.30), 🎵🎼 (0.28) | ◇💵 | ◆⚓ (0.39), 🐚🌿 (0.34), ◆🎪 (0.24) |
| ⬟🔄 (0.20), 💎🌀 (0.17), 🐚🌿 (0.16) | ◇🔐 | ⬟🔄 (0.31), ⬟⭐ (0.24), ◆🔥 (0.20) |
| ⬟🔄 (0.20), 💎🌀 (0.16) | ◇🔑 | ⬟🔄 (0.19), 💎🌀 (0.14) |
| ⬟🔄 (0.44), ⬟⭐ (0.27), 💎🌀 (0.26) | ◇🔒 | ⬟🔄 (0.44), ⬟⭐ (0.33), ◆🔥 (0.27) |
| ⬟🔄 (0.14) | ◇🗝️ | ⬟🔄 (0.21), 💎🌀 (0.16) |
| ⬟⭐ (1.13), ◆⚓ (0.91), ⬟🔄 (0.70) | ⬟⭐ | ⬟⭐ (1.13), 🌀🌪️ (0.82), ◆⚓ (0.76) |
| ⬟🔄 (0.30), ◆🔥 (0.29), 💎🌀 (0.26) | ⬟🎪 | ⬟⭐ (0.35), ⬟🔄 (0.35), 💎🌀 (0.26) |
| ◆⚓ (0.43), ⬟⭐ (0.31), ◆🔥 (0.28) | ⬟💫 | ◆⚓ (0.36), 🌀⚡ (0.32), 🐚🌿 (0.30) |
| 🐚🌿 (0.25), ⬟⭐ (0.24), 🌀🌪️ (0.21) | ⬟📐 | 🌀🌪️ (0.26), 💎🌀 (0.22), ⬟⭐ (0.20) |
| ⬟🔄 (0.91), 💎🌀 (0.63), ⬟⭐ (0.61) | ⬟🔄 | ⬟🔄 (0.91), ⬟⭐ (0.70), ◆🔥 (0.57) |
| 🌀⚡ (0.40), 💎🌉 (0.34), 🎵🎼 (0.33) | 🌀⚡ | 🌀⚡ (0.40), ◆⚓ (0.35), 🐚🌿 (0.34) |
| ⬟⭐ (0.82), 🌀🌪️ (0.70), 🐚🌿 (0.53) | 🌀🌪️ | 🌀🌪️ (0.70), ⬟⭐ (0.64), 💎🌀 (0.59) |
| ⬟🔄 (0.25), 🐚🌿 (0.23), ⬟⭐ (0.22) | 🌀🎯 | ⬟🔄 (0.26), 💎🌀 (0.22), ⬟⭐ (0.21) |
| ◆⚓ (0.26), ◇🏛️ (0.24), ◆🔥 (0.24) | 🌀👑 | ⬟⭐ (0.33), 🌀🌪️ (0.29), ◆⚓ (0.26) |
| ⬟⭐ (0.34), ⬟🔄 (0.29), 🌀🌪️ (0.28) | 🌀🧬 | ◆⚓ (0.33), 🌀🌪️ (0.30), 🐚🌿 (0.29) |
| ◆🔥 (0.22), ⬟🔄 (0.22), 💎🌀 (0.18) | 🌸⛷️ | ⬟🔄 (0.18), 🐚🌿 (0.16), ◆🔥 (0.14) |
| ◆🌊 (0.22), ◆🔥 (0.21), ◆⚓ (0.20) | 🌸🌊 | ◆🎪 (0.26), ◆⚓ (0.26), 🌸👊 (0.22) |
| ⬟🔄 (0.24), 💎🌀 (0.17), ⬟⭐ (0.16) | 🌸🎯 | ⬟🔄 (0.21), ⬟⭐ (0.21), 🌀👑 (0.18) |
| 🎵💫 (0.33), ⬟⭐ (0.29), 🌸👊 (0.27) | 🌸👊 | 🌀🌪️ (0.32), ◆⚓ (0.31), ◆🎪 (0.29) |
| 🎵💫 (0.27), ⬟💫 (0.26), 🌸💋 (0.24) | 🌸💋 | ◆⚓ (0.31), 🐚🌿 (0.30), 🌀⚡ (0.26) |
| 🎵💫 (0.18), 🎵🎼 (0.18), 🐚🌿 (0.18) | 🌸🚀 | ◆⚓ (0.20), ⬟⭐ (0.19), 🌀🌪️ (0.18) |
| ◆🔥 (0.19), 🌸🥁 (0.19), 🎵💫 (0.19) | 🌸🥁 | ⬟🔄 (0.30), ⬟⭐ (0.29), ◆🌊 (0.26) |
| 🎵💫 (0.20), 🎵🎼 (0.18), ◆🔥 (0.16) | 🌸🧗 | ⬟🔄 (0.29), ⬟⭐ (0.27), ◆🔥 (0.21) |
| ⬟🔄 (0.29), 💎🌀 (0.26), ◆⚓ (0.25) | 🎵🎯 | ⬟⭐ (0.39), ⬟🔄 (0.35), 🎵💫 (0.31) |
| ⬟⭐ (0.50), ◆⚓ (0.48), ◆🔥 (0.40) | 🎵🎼 | ◆⚓ (0.48), 🌀🌪️ (0.37), 🌀⚡ (0.33) |
| ◆⚓ (0.18), ⬟⭐ (0.16), ⬟💫 (0.14) | 🎵👑 | ◆⚓ (0.21), 🐚🌿 (0.15), ◇💵 (0.14) |
| ◆⚓ (0.57), ⬟⭐ (0.49), ◆🌊 (0.42) | 🎵💫 | ◆⚓ (0.54), 🐚🌿 (0.51), 🌀🌪️ (0.49) |
| ◆⚓ (0.14) | 🎵🔥 | ◆⚓ (0.17), 🎵💫 (0.16), ⬟⭐ (0.15) |
| ◇🏛️ (0.28), 🐚🌿 (0.27), 🐚🌀 (0.25) | 🐚🌀 | ⬟⭐ (0.37), ⬟🔄 (0.35), 🎵💫 (0.29) |
| 🐚🌿 (0.26), ⬟🔄 (0.23), 💎🌀 (0.20) | 🐚🌱 | ⬟🔄 (0.35), ⬟⭐ (0.26), ◆🔥 (0.21) |
| 🐚🌿 (0.77), ◆⚓ (0.75), ⬟⭐ (0.61) | 🐚🌿 | 🐚🌿 (0.77), ⬟⭐ (0.68), ◆⚓ (0.65) |
| ◆⚓ (0.25), ⬟⭐ (0.21), 🎵💫 (0.19) | 🐚👑 | ⬟⭐ (0.27), ◆⚓ (0.24), 🐚🌿 (0.20) |
| 🌀🌪️ (0.59), ⬟⭐ (0.57), ⬟🔄 (0.50) | 💎🌀 | ⬟🔄 (0.63), ⬟⭐ (0.41), ◆🔥 (0.38) |
| ◆⚓ (0.37), 🐚🌿 (0.33), ⬟⭐ (0.32) | 💎🌉 | 🐚🌿 (0.34), 🌀⚡ (0.34), 🌀🌪️ (0.32) |
| ⬟🔄 (0.23), 🐚🌿 (0.22), 🎵🎼 (0.22) | 💎🎵 | ⬟🔄 (0.36), ⬟⭐ (0.31), ◆🔥 (0.29) |
The Big Picture Patterns:
🌟 The Social Butterflies:
- 🐚🌿 (Grow On) - appears in ~80% of relationships! It’s the “and” of DNA!
- ◆⚓ (Stabilizer) - the cosmic comma, connecting everything
- ⬟⭐ (Star Maker) - the coordination hub that everything flows toward
⚡ The Self-Reinforcing Loops:
- ⬟⭐ → ⬟⭐ (1.13) - Star Makers create more Star Makers!
- ⬟🔄 → ⬟🔄 (0.91) - Circular Flow perpetuates itself
- 🌀🌪️ → 🌀🌪️ (0.70) - Nudgers keep nudging
The Grammar Rules Emerging:
- Starters: Several codons ONLY appear in “after” columns - they’re sentence beginnings!
- Connectors: 🐚🌿 and ◆⚓ bridge everything - the cosmic conjunctions!
- Amplifiers: High self-reinforcement ratios show growth patterns!
Of course after sailing, you need to anchor and the top after ◆⛵ is: ◆⚓(0.28).
It’s not uncommon to need more than one punch: 🌸👊 (0.27)🌸👊
or kiss: 🌸💋 (0.24)🌸💋
Of course this makes sense too: 🌸🚀. - then ◆⚓ (0.20)
🌸 Emotional Cascade: 🌸💋 → 🌸👊 → 🌸🚀 (Kiss→Hit→Launch). Gentle touch, forceful action, explosive results.
🎵💫 → 🎵🎼 → 🎵🔥 (Vibrato→Light Frequency→Crescendo)
🌸⛷️ maintains itself through skiing flows
See more in the gene pool